
CHLAMYDOMONAS FLAGELLA AND PHOTOBEHAVIORS

ここでは、
クラミドモナス Chlamydomonas reinhardtii
クラミドモナス(学名: Chlamydomonas reinhardii; 和名: コナミドリムシ)は、淡水に棲む単細胞の緑藻です。
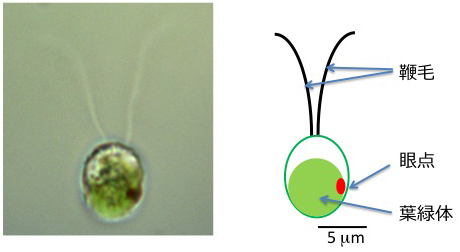
クラミドモナスの顕微鏡写真と模式図。長さ約12 μmの2本の鞭毛を平泳ぎのように動かして水中を泳ぎます。眼点で光を感受して、光合成を行うのに最適な光の環境へと移動します。
クラミドモナスは実験生物として以下のメリットを持ち、鞭毛・繊毛、光合成、有性生殖など、さまざまな研究分野でモデル生物として使われています。特に鞭毛・繊毛の研究では、分野をリードする立場にあります。
- 通常は一倍体で無性的に増殖するため、ミュータントの作成が容易。
- 2つの性をもち、有性生殖を行うため、古典遺伝学的手法による遺伝子解析(四分子解析)が行える。
- 鞭毛の単離・精製法が確立されている。
- 外来遺伝子導入法が確立されている。
- 培養スケールが容易に変えられる。(寒天培地上での少量培養から、10リットル以上の大量液体培養まで。)
- 全ゲノム解読が終了しており、ゲノム、EST、プロテオーム等の有用なデータベースが公開されている。
■クラミドモナスの運動の様子(動画ファイルが別ウインドウで開きます。)
- 暗視野顕微鏡で観察した遊泳の様子。(2.8 MB)(鞭毛を約60 Hzで動かし、約120 μm/secで泳ぎます。)
- 暗視野顕微鏡+高速度カメラでの細胞観察。(1.3 MB)(2枚のガラスで挟んだ細胞を撮影。1/40倍速で再生。)
真核生物の鞭毛・繊毛
真核生物の鞭毛・繊毛は、細胞に生えた毛のような細胞小器官です。(1細胞から数本程度生えている長いものを「鞭毛」、たくさん生えている短いものを「繊毛」と呼ぶ習慣がありますが、両者の構造・運動機構は本質的に同じです。)
真核生物の鞭毛・繊毛の横断面は、クラミドモナスのような原生生物からヒトまで共通した「9+2構造」をもちます。(9組の二連周辺微小管が2本の中心対微小管を取り囲むためにそう呼ばれます。)
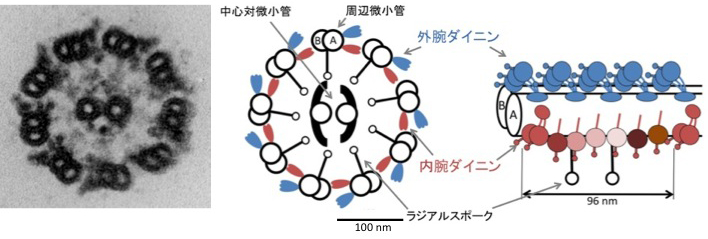
図の左はクラミドモナスの鞭毛の横断面の電顕像、真ん中が模式図、右は1組の周辺微小管を長さ方向に見た模式図です。
周辺微小管のA小管の上に規則正しく結合したモータータンパク質「ダイニン」が、向かい側のB小管に対して、ATPの加水分解のエネルギーを利用して滑りを起こします。これが鞭毛運動の原動力です。
ダイニンは外腕と内腕に大別されます。外腕ダイニンは1種類が24 nm周期で1列に、内腕ダイニンは7種類がそれぞれの位置に複雑に並んで96 nm周期で結合しています。(他にも、鞭毛の根元だけに局在する特殊な内腕ダイニンの存在が知られています。)滑り活性などの個性の異なる複数のダイニンが、それぞれ決まった位置に周期的に並ぶことによって、鞭毛は波打ち運動を伝播することができると考えられています。
ダイニンが正しい位置で正しく機能するメカニズムの理解は、鞭毛の運動メカニズムを理解する上で重要です。私達は、鞭毛運動の出力の3分の2を生み出す外腕ダイニンに着目しています。外腕ダイニンは約20のサブユニットから成る複合体であり、リン酸化、カルシウム、さらにレドックス(酸化・還元)といったさまざまな化学シグナルを受け取ることが知られています。私達は、「外腕ダイニンが決まった位置に24 nm周期で配列するしくみ」「外腕ダイニンの活性が(特にレドックス・シグナルによって)調節されるしくみ」を研究しています。
■関連論文
-
Wakabayashi K, Takada S, Witman GB, Kamiya R.
Transport and arrangement of the outer-dynein-arm docking complex in the flagella of Chlamydomonas mutants that lack outer dynein arms.
Cell Motil Cytoskeleton. 2001 Apr;48(4):277-86. -
Takada S, Wilkerson CG, Wakabayashi K, Kamiya R, Witman GB.
The outer dynein arm-docking complex: composition and characterization of a subunit (oda1) necessary for outer arm assembly.
Mol Biol Cell. 2002 Mar;13(3):1015-29. -
Wakabayashi K, King SM.
Modulation of Chlamydomonas reinhardtii flagellar motility by redox poise.
J Cell Biol. 2006 Jun 5;173(5):743-54.
(Faculty of 1000推薦論文) -
Wakabayashi K, Ide T, Kamiya R.
Calcium-dependent flagellar motility activation in Chlamydomonas reinhardtii in response to mechanical agitation.
Cell Motil Cytoskeleton.2009 Sep;66(9):736-42.
-
Wakabayashi K.
Regulation of Axonemal Outer-Arm Dyneins in Cilia
Dyneins: Structure, Biology and Disease.Chapter 10 (297-311) 2011 Elsevier Inc.
鞭毛・繊毛というと、「微生物の運動装置」という印象を持つ人が多いでしょう。しかし、ヒトのさまざまな器官(脳、気管上皮、男性の精子、女性の輸卵管など)に鞭毛・繊毛が生えています。それらが生えない・動かないことによって起こる「原発性繊毛運動不全症(primary ciliary dyskinesia, PCD)」の患者さんが1万人に1人の割合で存在します。(PCDは慢性副鼻腔炎、気管支拡張症、水頭症、男性不妊症、内臓逆位などの症状が現れます。)クラミドモナスを使った鞭毛研究は、このPCDの理解に大きく貢献してきました。というのは、鞭毛・繊毛を生やす・動かすのに必要な遺伝子はヒトとクラミドモナスでほとんど共通だからです。いま、クラミドモナスの鞭毛研究は医学的にも注目を集めています。
クラミドモナスの光行動
クラミドモナス眼点の光受容体「チャネル型ロドプシン」は、光を受容するとカチオンを流入する「イオンチャネル」としての機能を持ちます。光受容後、膜の興奮によって鞭毛にカルシウムイオンが流入し、光の強さに応じて(流入するカルシウムの量に応じて)2つの光行動を示します。
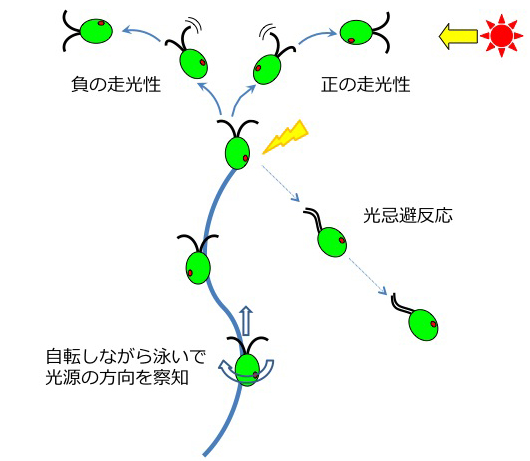
2つの光行動の概略。強い光を浴びると、鞭毛の波形を変換して遊泳方向を逆転させる「光忌避反応」を示します。弱い光を浴びると、2本の鞭毛を打つバランスを変化させて「正または負の走光性」を示します。
【1】光忌避反応/光驚動反応 (photophobic response, photoshock response)
カメラのフラッシュのように急激に強い光を浴びたとき、カルシウムイオンが10-4 M 程度鞭毛に流入すると考えられています。すると鞭毛は打ち方を通常の「平泳ぎ」(繊毛型、非対称型波形などと呼びます)から「ドルフィンキック」(鞭毛型、対称型波形などと呼びます)のような動かし方に変換して、約1秒間後ろ向きに泳ぎます。これを光忌避反応/光驚動反応と呼びます。
- クラミドモナスの光忌避反応。(1.6 MB)(クラミドモナス眼点の光受容体は青?緑色の光を感受し、赤い光を感じることはできません。この動画では、赤いフィルターを急に外すことによって光刺激を与えています。「びっくり」した細胞は約1秒間後ろ向きに泳ぎ、やがて前向き泳ぎに戻ります。)
【2】走光性/光走性(phototaxis, phototactic response)
弱い光を感受すると、カルシウムイオンが10-7 M程度鞭毛に流入すると考えられています。クラミドモナスの2本の鞭毛は眼点に近いか遠いかで区別され、それぞれカルシウム感受性が異なる事が知られています。その性質を利用して舵取りを行い、クラミドモナスは光源に向かう正の走光性と、逆に光源から逃げる負の走光性を示します。(動物学の分野では走光性(そうこうせい)、植物学の分野では光走性(ひかりそうせい)と呼ぶ習慣があるようです。)
- クラミドモナスの正の走光性。(7.6 MB)(最初はランダムな方向に泳いでいますが、カウンターが5秒になったところで左から、15秒になったところで上から緑色の光を当てています。クラミドモナスは光の照射方向変化に応じて迅速に遊泳方向を変えます。)
【走光性のレドックス調節】
走光性の正・負の切り替えは、クラミドモナスが光合成のために最適な光環境を維持するために重要であると考えられます。しかし、その切り替えメカニズムはよくわかっていませんでした。私達は、細胞内のレドックス(酸化・還元)状態の変化がそれを切り替えており、酸化的になると正、還元的になると負の走光性を示すということを見出しました。
-
■同じ培養液から採取した細胞が、レドックス薬剤によって走光性の正負を逆転させた様子。ともに右側から光を照射しています。
- 正の走光性を示す細胞(無処理)(1.2 MB)
- 負の走光性を示す細胞(膜透過性 活性酸素消去剤処理)(1.2 MB)
細胞はさまざまシステムによって「レドックス恒常性」を維持し、タンパク質・脂質等の酸化や過剰な還元を防いでいます。レドックスによる走光性の正負切り換えは、「光強度変化→光合成活性変化→細胞内レドックス状態変化→走光性の正負の切り換え→光強度変化」というフィードバックによってレドックス恒常性維持に貢献していると考えられます。現在私達は、クラミドモナスがどのようにして細胞内のレドックス状態変化を察知し、その情報をどのようにして鞭毛打のバランス変化につなげているのか、その分子メカニズムを研究しています。
■関連論文
-
Wakabayashi K, Misawa Y, Mochiji S, Kamiya R.
Reduction-oxidation poise regulates the sign of phototaxis in Chlamydomonas reinhardtii
Proc Natl Acad Sci U S A. 2011 Jul 5;108(27):11280-4. Epub 2011 Jun 20.
(Faculty of 1000推薦論文) -
Mochiji S, Wakabayashi K.
Redox regulation of phototactic migration in the green alga Chlamydomonas reinhardtii and its possible application.
Commun Integr Biol. 2012 Mar 1;5(2):196-8.