
Latest Research
- 2016.12.16
- Tanaka-Yoshida Group
Function of plant hormone abscisic acid in a primitive red alga
Plant hormones are chemical compounds that are induced by environmental changes and stresses. Plant hormones play important roles in various physiological responses such as tissue differentiation, growth, and morphogenesis in land plants. Among them, abscisic acid (ABA) induces a stress response, closing of stomata, seed dormancy and so on. It plays an extremely important role in the environmental adaptation of plants1. ABA has been found in prokaryotic cyanobacteria and photosynthetic unicellular algae while function in these organisms remains poorly understood. Hence, it is unclear when ABA was established as a plant hormone during the plant evolution. To approach this issue, we focused on Cyanidioschyzon merolae as a most primitive plant-type organism. C. merolae is a unicellular red alga living in a sulfur acidic hot spring. It has excellent advantages as a model organism, which contains one plastid, one mitochondrion, and one nucleus as the simplified model system, and all genome sequences were completely determined2.
To understand the establishment of origin of the ABA hormone signaling, we analyzed the ABA function in C. merolae. ABA synthesis genes were identified based on C. merolae genome information. Then, we verified whether ABA is actually detected from C. merolae cells. In land plant, ABA is detected even in the normal growth condition. However, ABA was not detected under normal growth condition in C. merolae, and detected only under high salt stress condition (Table 1). To examine the function of ABA, we dosed extracellular ABA into C. merolae culture. ABA was found to inhibit the cell cycle at the G1 to S phase transition. In a previous study, we showed that tetrapyrrole molecules play critical roles in the cell cycle progression of C. merolae3. In plant cells as C. merolae, tetrapyrrole molecules are present as chlorophyll, heme and their intermediates. It was found that intracellular free (unfettered form) heme decreased under the ABA dosed condition (Figure 1A). It was also found that ABA regulates the free heme accumulation throught the expression of heme scavenger protein TSPO in C. merolae. It was also reported in land plants that TSPO is induced by ABA and regulates free heme accumulation4. These observations suggested that regulation of free heme accumulation by ABA is conserved from primitive algae to land plants. Furthermore, we found that ABA induced changes of free heme accumulation results in the inhibition of cell cycle G1/S transition in C. merolae5, 6 (Figure 1B).
Table 1. ABA accumulation in C. merolae cells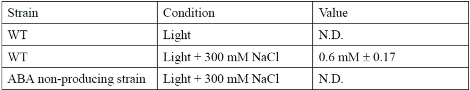
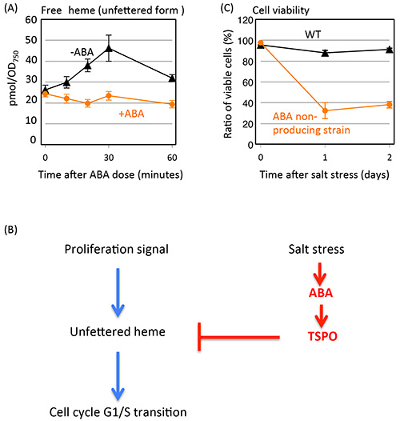
Figure 1. Changes of amounts of unfettered heme, model of ABA function in C. merolae and salt tolerance in ABA non-producing strain.
Why does ABA need to arrest the cell cycle ? Since ABA accumulates under salt stressed condition, we supposed that it would be related to salt stress tolerance. To verify this, we constructed an ABA non-producing strain and compared its growth and viability with wild type cells under salt stressed condition. While wild type cells retained viability even under high salt condition, ABA non-producing strain cells showed a significant loss of viability under the same condition (Figure 1C). Salt stress is known to generate reactive oxygen spices (ROS) in vivo. Cells are vulnerable to ROS during the DNA replication and division phases of the cell cycle. Thus, our results indicated that ABA protects cells from salt stress by arresting at the G1 phase.
In this study, we showed that a primitive alga C. merolae has an ABA-dependent stress resistant system (Figure 2). It is suggested that ABA related stress responsive system was established very early in the evolution of photosynthetic eukaryotes.
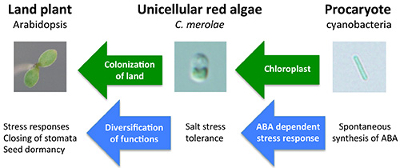
Figure 2. Model of ABA evolution.
References
1. Hauser, F., Waadt, R. and Schroeder, J.I. (2011) Evolution of abscisic acid synthesis and signaling mechanisms. Curr. Biol. 21: R346-R355.
2. Kobayashi, Y., Ohnuma, M., Kuroiwa, T., Tanaka, K. and Hanaoka, M. (2010) The basics of cultivation and molecular genetic analysis of the unicellular red alga Cyanidioschyzon merolae. Endocytobiosis Cell Res. 20: 53-61.
3. Kobayashi, Y., Kanesaki, Y., Tanaka, A., Kuroiwa, H., Kuroiwa, T. and Tanaka, K. (2009) Tetrapyrrole signal as a cell-cycle coordinator from organelle to nuclear DNA replication in plant cells. Proc. Natl Acad. Sci. USA 106: 803-807.
4. Vanhee, C., Zapotoczny, G., Masquelier, D., Ghislain, M. and Batoko, H. (2011) The Arabidopsis multistress regulator TSPO is a heme binding membrane protein and a potential scavenger of porphyrins via an autophagy-dependent degradation mechanism. Plant Cell 23: 30 785-805.
5. Kobayashi, Y., Ando, H., Hanaoka, M. and Tanaka, K. (2016) Abscisic acid participates in the control of cell-cycle initiation through heme homeostasis in the unicellular red alga Cyanidioschyzon merolae. Plant Cell Physiol. 57, 953-960.
6. Kobayashi, Y. and Tanaka, K. (2016) Transcriptional regulation of tetrapyrrole biosynthetic genes explains abscisic acid-induced heme accumulation in the unicellular red alga Cyanidioschyzon merolae. Front. Plant Sci. 29 (http://dx.doi.org/10.3389/fpls.2016.01300)
