
RESEARCH_PROJECTS
緑藻の光反応行動 【1】クラミドモナス
クラミドモナスの眼点付近に局在する光受容体「チャネルロドプシン」は、光を受容すると陽イオンを流入します。その後細胞膜・鞭毛膜が興奮して鞭毛にカルシウムイオンが流入し、光の強さに応じて(流入するカルシウムの量に応じて)2つの光行動を示すと考えられています。
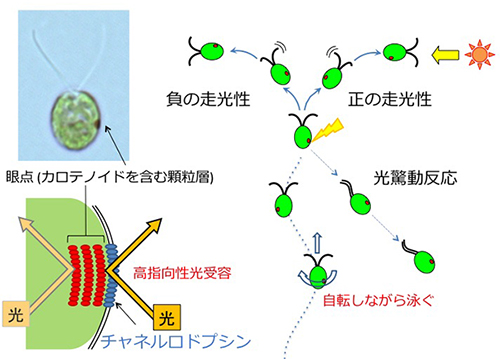
2つの光行動の概略図。
眼点の実態は、カロテノイド色素を豊富に含んだ顆粒が層状に配列したものです。この構造が光を反射するため、その直上の細胞膜に存在するチャネルロドプシンは、細胞の外側からきた光しか感受しません。また、二本の鞭毛の打面がずれていることから、細胞は自転しながら泳いでいます。
この「高指向性光受容」「自転遊泳」の2つの要素により、クラミドモナスは光の入射方向を正確に感知すると考えられています。
カメラのフラッシュのような急激な強い光の照射に対しては光驚動反応で遊泳方向を逆転させます。連続的な弱い光の照射に対しては正または負の走光性を示します。
■光驚動反応/光忌避反応 (Photoshock response/Photophobic response)
急激に強い光を浴びたとき、カルシウムが10-4 M程度鞭毛に流入すると考えられています。すると鞭毛は打ち方を通常の「平泳ぎ」(繊毛型、非対称型波形などと呼びます)から「ドルフィンキック」(鞭毛型、対称型波形などと呼びます)のような動かし方に変換して、1秒間程度後ろ向きに泳ぎます。これを光驚動反応(あるいは光忌避反応)と呼びます。
-
 クラミドモナスの光驚動反応。(2.2 MB)(高速度カメラで撮影し、1/30倍速で再生しています。カウンターが70になったところでカメラのフラッシュを焚きました。その後しばらくの間、後ろ向きに泳ぎます。)
クラミドモナスの光驚動反応。(2.2 MB)(高速度カメラで撮影し、1/30倍速で再生しています。カウンターが70になったところでカメラのフラッシュを焚きました。その後しばらくの間、後ろ向きに泳ぎます。)
■走光性/光走性(Phototaxis)
弱い光を感受すると、カルシウムが10-7 M程度鞭毛に流入すると考えられています。クラミドモナスの2本の鞭毛は眼点に近いか遠いかで区別され、それぞれカルシウム感受性が異なる事が知られています。その性質を利用して舵取りを行い、クラミドモナスは光源の方向に向かい正の走光性と、逆に光源から逃げる負の走光性を示します。(動物学の分野では走光性(そうこうせい)、植物学の分野では光走性(ひかりそうせい)と呼ぶ習慣があるようです。)
-
 クラミドモナスの正の走光性。(7.6 MB)(最初はランダムな方向に泳いでいますが、カウンターが5秒になったところで左から、15秒になったところで上から緑色の光を当てています。クラミドモナスは光の照射方向変化に応じて迅速に遊泳方向を変えます。)
クラミドモナスの正の走光性。(7.6 MB)(最初はランダムな方向に泳いでいますが、カウンターが5秒になったところで左から、15秒になったところで上から緑色の光を当てています。クラミドモナスは光の照射方向変化に応じて迅速に遊泳方向を変えます。)
*
走光性の正・負の切り替えは、クラミドモナスが光合成のために最適な光環境を維持するために重要であると考えられます。しかし、その切り替えメカニズムはよくわかっていませんでした。私達は、細胞内のレドックス(酸化・還元)状態の変化がそれを切り替えており、酸化的になると正、還元的になると負の走光性を示すということを見出しました。
下の動画は、同じ培養液から採取した細胞の片方にだけ活性酸素消去処理をしたものです。ともに右側から光を照射していますが、無処理細胞は右へ(正の走光性)、活性酸素消去処理をした細胞は左へ(負の走光性)泳ぎます。
 正の走光性を示す細胞(無処理)(1.2 MB)
正の走光性を示す細胞(無処理)(1.2 MB) 負の走光性を示す細胞(膜透過性 活性酸素消去剤処理)(1.2 MB)
負の走光性を示す細胞(膜透過性 活性酸素消去剤処理)(1.2 MB)現在私達は、クラミドモナスがどのようにして細胞内のレドックス状態変化を察知し、その情報をどのようにして鞭毛打のバランス変化につなげているのか、その分子メカニズムを研究しています。その過程で、いつも野生株と逆方向に泳ぐ「あまのじゃく」変異株を単離しました。この株は眼点のカロテノイド色素の生合成経路を欠損しています。そのため、細胞のレンズ効果により、光源方向を勘違いしてしまうのです。

あまのじゃく変異株lts1-211の表現型
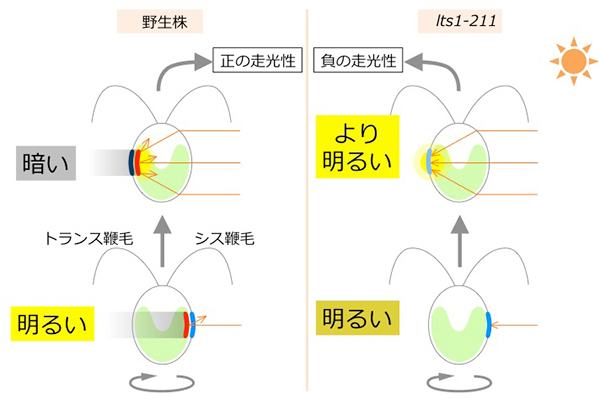
lts1-211の「あまのじゃく」は細胞レンズ効果で説明できる

細胞レンズ効果の証明方法。顕微鏡の視野絞りに「P」と印刷したOHPシートを置く。そしてフォーカスを細胞から徐々に上に上げると…(下の動画をご覧ください)
 細胞レンズ効果の証明(6.5 MB)
細胞レンズ効果の証明(6.5 MB)■関連する最近の研究成果
-
Mochiji S, Wakabayashi K.

Redox regulation of phototactic migration in the green alga Chlamydomonas reinhardtii and its possible application.
Commun Integr Biol. 2012 Mar 1;5(2):196-8.
-
Wakabayashi K, Misawa Y, Mochiji S, Kamiya R.
Reduction-oxidation poise regulates the sign of phototaxis in Chlamydomonas reinhardtii
Proc Natl Acad Sci U S A. 2011 Jul 5;108(27):11280-4. Epub 2011 Jun 20.
(Rated as "Recommended" in "Faculty of 1000") -
Ide T, Mochiji S, Ueki N, Yamaguchi K, Shigenobu S, Hirono M, Wakabayashi K.
Identification of the agg1 mutation responsible for negative phototaxis in a "wild-type" strain of Chlamydomonas reinhardtii.
Biochem Biophys Rep. 2016 Jul 20;7:379-385. doi: 10.1016/j.bbrep.2016.07.016. eCollection 2016 Sep. -
Ueki N, Ide T, Mochiji S, Kobayashi Y, Tokutsu R, Ohnishi N, Yamaguchi K, Shigenobu S, Tanaka K, Minagawa J, Hisabori T, Hirono M, Wakabayashi K.
Eyespot-dependent determination of the phototactic sign in Chlamydomonas reinhardtii
Proc Natl Acad Sci U S A. 2016 May 10;113(19):5299-304. doi: 10.1073/pnas.1525538113. Epub 2016 Apr 27.
(平成28年度手島精一記念研究賞研究論文賞受賞)
緑藻の光反応行動 【2】ボルボックス
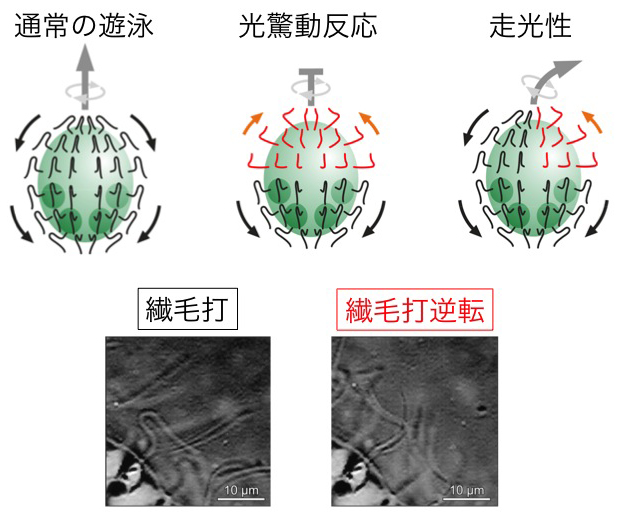
ボルボックスの細胞一つ一つは、鞭毛を二本と眼点を一つ持ち、クラミドモナスとよく似ています。しかし、ボルボックスは決して「クラミドモナスが集まったもの」ではありません。ボルボックスの細胞は、二本の鞭毛を平泳ぎではなく、同じ方向へと打っています。数千の細胞から生える鞭毛が全て、個体の前から後ろへと打つことで、大きなボルボックスが全体として前へと進むことができるのです。
■光驚動反応
ボルボックスも光驚動反応を示します。フラッシュのような光が当たると、光に敏感な前の方の細胞が全て鞭毛の向きを逆転させます。これによって、大きなボルボックス全体にブレーキがかかり、ボルボックス個体はぴたっと止まります。
■走光性
ボルボックスの走光性は非常に巧妙なしくみで行われます。一方向から光が当たるとき、個体の自転によって細胞が光側に来るときに明るい光を感じます。そのとき鞭毛の打つ向きが逆転し、すぐに元に戻ります。その結果、常に光側の細胞の鞭毛だけが逆向きに打つことになり、ボルボックス全体の進む力がアンバランスになって光の方へと向きを変えるのです。「明るくなったら逆に打つ」というたった一つのルールによって、細胞同士が連絡し合わなくても、数千の細胞が協調した行動をすることができるのです。
ボルボックスは、クラミドモナスのような単細胞性の祖先生物が進化の過程で多細胞化したものと考えられています。そのとき、一つの細胞としてではなく一つの多細胞生物として行動するしくみをどのように獲得したのでしょうか?最近私たちは、ボルボックス個体をまるごと界面活性剤処理し、ここにATPを添加することで「死んでいるのに泳ぐ」ゾンビ・ボルボックス法を開発しました。これにより、ボルボックス鞭毛の制御シグナルの候補であるカルシウムイオンの効果を直接調べることができます。この実験により、個体の前端から後端にかけて鞭毛のカルシウムに対する応答性に勾配があることが初めてわかりました。前端部はカルシウムイオン添加によって打つ方向をほぼ180度逆転し、赤道面付近は90度回転し、後端部付近はほとんど運動を変化させません。ボルボックスは、光を受容した際に生じるカルシウムイオン流入に応じた鞭毛運動変化に勾配をつけることによって、大きな体で迅速に光行動を示すことを可能にしているのです。
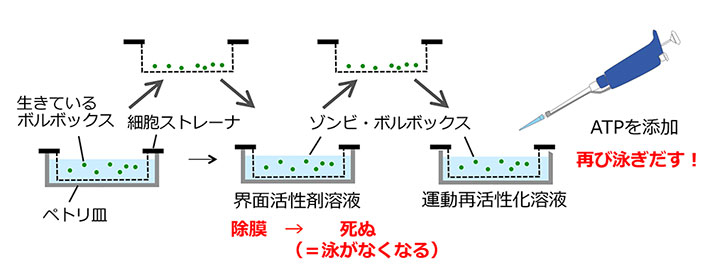 ゾンビ・ボルボックス法の概要
ゾンビ・ボルボックス法の概要
 ゾンビ・ボルボックス実験(6.5 MB)
ゾンビ・ボルボックス実験(6.5 MB)
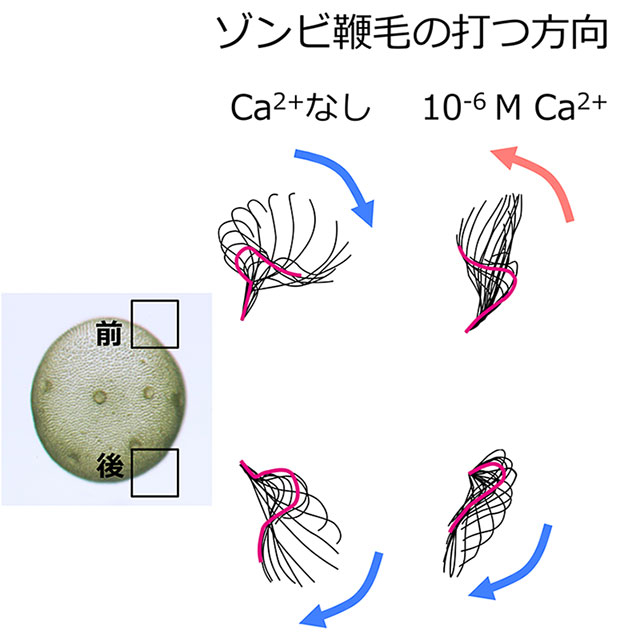
ゾンビ・ボルボックス法でわかったボルボックス鞭毛のカルシウムに対する応答性の勾配
■関連する最近の研究成果
-
Ueki N, Matsunaga S, Inouye I, Hallmann A.
How 5000 independent rowers coordinate their strokes in order to row into the sunlight: phototaxis in the multicellular green alga Volvox.
BMC Biol.2010 Jul 27;8:103. doi: 10.1186/1741-7007-8-103.
-
Ueki N, Wakabayashi K.
Detergent-extracted Volvox model exhibits an anterior-posterior gradient in flagellar Ca2+ sensitivity.
Proc Natl Acad Sci U S A. 2018 Jan 30;115(5):E1061-E1068. doi: 10.1073/pnas.1715489115. Epub 2018 Jan 8.